
Intensieve training activeert de eiwitsynthese, maar alleen als de juiste voeding er is om deze te ondersteunen. Als je het artikel dat John Meadows en ik schreven met de titel Maximize Protein Synthesis hebt gelezen, of als je iets hebt gelezen dat T Nation al jaren over het onderwerp zegt, ben je waarschijnlijk bekend met het concept van 'het anabole venster' en het belang ervan. van peri-workout voeding.
Wat er in de uren na de training op cellulair niveau gebeurt, is voorspellend voor winst op de lange termijn. Profiteer van dit "anabole venster" en je zult groeien als nooit tevoren. Mis het consequent, en, nou ja, veel geluk.
De juiste macronutriënten op het juiste moment krijgen is de sleutel, maar macro's zijn slechts een deel van het grote geheel. Een belangrijk maar vaak over het hoofd gezien aspect van spiereiwitsynthese is het celvolume. Celvolume is niet alleen cosmetisch; het is de belangrijkste motor van het transport van aminozuren en werkt ook achter de schermen om de eiwitsynthese in te schakelen en de afbraak van eiwitten te onderdrukken.
Een volle / volumineuze spier is een anabole spier. Hoewel we al meer dan 20 jaar weten dat celzwelling de afbraak van eiwitten remt en de eiwitsynthese in bepaalde cellen stimuleert (1-3), was tot voor kort het mechanisme dat celvolume koppelde aan eiwitsynthese een mysterie.
Wat we nu weten is dat de eiwitsynthese wordt gecontroleerd door het enzym mTOR, dat wordt geactiveerd door mechanische stress, groeifactoren en leucine.
Hoewel deze alle drie belangrijk zijn voor de trainingsprikkel, is mTOR-signalering ook afhankelijk van het celvolume.(4) Dit is vooral belangrijk in skeletspieren, waar celvolumisatie de glycogeen- en eiwitsynthese activeert en de eiwitafbraak remt.(5, 6)
De wetenschappelijke doorbraak die leidde tot het verband tussen celvolume en eiwitsynthese vond plaats in 2005, toen een groep wetenschappers ontdekte dat er meer nodig is dan alleen leucine om mTOR te activeren - glutamine is ook vereist.(7)
Dit kwam als een verrassing. Hoewel glutamine wordt beschouwd als een "voorwaardelijk essentieel" aminozuur dat de eiwitafbraak beperkt tijdens ernstig trauma of stress, was het nooit in verband gebracht met mTOR-activering.
Glutamine was nodig voor de opname van leucine en celvolumisatie, die beide nodig zijn om de eiwitsynthese in te schakelen. De auteurs toonden aan dat cellulaire glutamine uitputting resulteert niet alleen in een vermindering van het celvolume, maar vermindert ook het vermogen van leucine om de eiwitsynthese te activeren.(7)
Deze ontdekking was enorm, omdat het een directe link bood tussen glutamine, celvolume en eiwitsynthese. Voor het eerst werd aangetoond dat glutamine nodig is voor de activering van eiwitsynthese door leucine.
Hoewel deze studie suggereerde dat glutamine een zeer belangrijk stuk is van de puzzel die celvolume en eiwitsynthese met elkaar verbindt, werd het exacte mechanisme pas in 2009 uitgewerkt, toen Nicklin et al. ontdekte dat glutamine-export gekoppeld is aan leucine-import en mTOR-activering.(8)
Om leucine in de cel te krijgen, is er een eerste periode van ‘glutamine’.“Dit trekt ook water aan, waardoor het celvolume toeneemt. Na de fase van het laden van glutamine wordt glutamine uit de cel geëxporteerd in ruil voor de invoer van leucine.
Nicklin et al. ontdekte ook dat cellulaire glutaminespiegels snelheidsbeperkend zijn voor de activering van eiwitsynthese door leucine. Wanneer cellen gelijktijdig werden behandeld met glutamine en een EAA-mengsel dat leucine bevat, werd de activering van de eiwitsynthese 60 minuten vertraagd. Toen deze zelfde cellen glutamine 'voorgeladen' waren, werd de eiwitsynthese ingeschakeld binnen 1-2 minuten nadat de cellen leucine hadden gekregen.
Dit resultaat was belangrijk omdat het de vertragingstijd verklaarde voor de activering van eiwitsynthese door leucine in dit experimentele model.
Deze resultaten werpen eindelijk enig licht op de cellulaire machinerie die het transport van aminozuren reguleert, en hoe deze is gekoppeld aan de controle van de eiwitsynthese.
Dit werk moet echter met enige voorzichtigheid worden geïnterpreteerd. Een belangrijk voorbehoud bij deze onderzoeken was dat ze in vitro werden uitgevoerd (i.e. in celculturen) waar regulering of eiwitsynthese veel eenvoudiger is. Spiercellen zijn in staat om naar behoefte glutamine te maken uit andere aminozuren, en "glutamine-uitputting" in dit celkweekmodel is niet representatief voor meer fysiologische situaties in vivo.
Hoge eiwitsynthesesnelheden kunnen niet voor onbepaalde tijd worden gehandhaafd in spierweefsel met of zonder glutaminesuppletie. Glutamine kan echter worden gebruikt om de eiwitsynthese strategisch te ondersteunen door het celvolume te optimaliseren tijdens de periode na de training.

Cellen zijn erg druk en er zijn veel membraangebonden ionenkanalen en transporteiwitten die het verkeer naar en uit de cel reguleren. Er zijn in het bijzonder twee klassen van aminozuurtransporters die hier van belang zijn: de aminozuurtransporters 'System L' en 'System A' zijn het nauwst verbonden met mTOR-signalering en eiwitsynthese.(8-10)
De activiteit van System A- en System L-transporters is gekoppeld, waardoor leucine en de andere BCAA's in de cel kunnen worden opgenomen.(11) Systeem L-transporteurs zijn verantwoordelijk voor de instroom van leucine en de andere BCAA's in ruil voor de uitstroom van glutamine.
Systeem A-transporteurs werken echter via een ander mechanisme, waarbij glutamine wordt gekoppeld aan natriumopname.(12, 13) De koppeling tussen natriumopname en System L / System A-aminozuurtransporters wordt Tertiair Actief Transport (afgekort TAT) genoemd. Het is TAT dat uiteindelijk leucine in de cel drijft, wat leidt tot mTOR-activering en eiwitsynthese.(11)
Je kunt zien hoe TAT werkt in de onderstaande afbeelding:
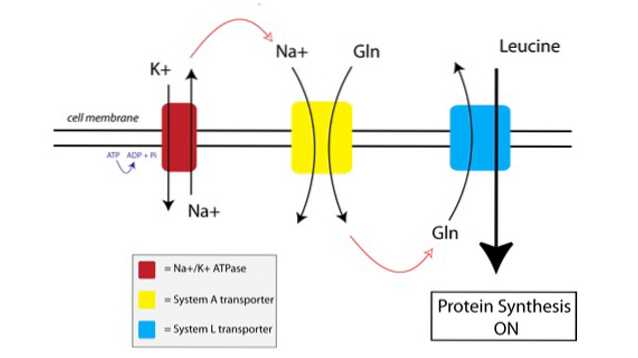
Ten eerste gebruikt een membraangebonden pomp, de natrium-kalium-ATPase-pomp (Na + / K + ATPase, rood in de bovenstaande afbeelding), energie van ATP om natrium buiten de cel te verplaatsen, tegen de concentratiegradiënt in.
De verhoogde natriumconcentratie buiten de cel is gekoppeld aan de invoer van glutamine door de System A-transporter (geel in de figuur). De instroom van glutamine en natrium in de cel trekt ook extra water aan, waardoor de cel opzwelt. Dit brengt de cel in een anabole toestand, waardoor de eiwitsynthese-machinerie gereed wordt gemaakt voor activering.
Wanneer glutamine zich opbouwt tot voldoende hoge niveaus in de cel, worden de System L-transporters (blauw in de figuur) geactiveerd, die glutamine naar buiten de cel transporteren in ruil voor opname van leucine. Het binnendringen van leucine in de cel is de trigger voor eiwitsynthese.
Hoewel dit tot nu toe een geweldige biochemieles is geweest, is de ontdekking van TAT niet alleen belangrijk voor celbiologen. Nu we weten hoe celvolume is gekoppeld aan aminozuurtransport en eiwitsynthese, kunnen we verschillende voedingsstrategieën ontwerpen om het proces te maximaliseren wanneer het telt - tijdens de kritieke periode na de training.
Eiwitsynthese is volledig afhankelijk van celhydratatie - als je zelfs een beetje uitgedroogd bent, is het vermogen om te herstellen van een intensieve training volledig aangetast. Voldoende water krijgen is hier een goed idee, maar water alleen is niet voldoende.
Elektrolyten zoals natrium, kalium, chloride en fosfaat functioneren ook als 'osmolieten' omdat ze water in de cel trekken. Na een intensieve trainingssessie hebben we water, aminozuren en elektrolyten nodig om het celvolumeproces dat de eiwitsynthese stimuleert te maximaliseren.
Natrium, magnesium, calcium, kalium, fosfaat en chloride (om er maar een paar te noemen) zijn hier allemaal belangrijk. Op een minimaal niveau (tenzij u dit doet op advies van uw arts), schroom niet om natrium vóór of na de training te vermijden. Als je natriumarm bent, zal je pomp van training bijna niet bestaan en is natrium nodig voor de opname van glutamine.
Om het giswerk te elimineren, zijn Surge® Workout Fuel en Plazma ™ ontworpen met ideale elektrolytverhoudingen om het celvolume en de eiwitsynthese te ondersteunen.
Opname van glutamine in de cel veroorzaakt celvolumisatie, waardoor spiercellen klaar zijn voor eiwitsynthese. Zoals gezegd is een volle / volumineuze spier een anabole spier. Naast het stimuleren van aminozuurtransport, verhoogt celvolumisatie ook de glycogeensynthese en remt het de afbraak van eiwitten.(4-6)
Eiwitsynthese wordt onderdrukt door de uitputting van glutamine, wat enorme gevolgen heeft voor hard trainende atleten. Na een intensieve trainingssessie wordt een ontstekingsreactie opgebouwd, waardoor immuuncellen in geslagen spierweefsel terecht kunnen komen om het herstel- / wederopbouwproces te starten.(14)
Glutamine wordt zo snel opgenomen door immuuncellen dat het wordt beschouwd als de “brandstof van het immuunsysteem”.”(15) Het is niet verrassend dat is aangetoond dat intensieve training plasmaglutamine-uitputting veroorzaakt.(16-18)
Om deze reden nemen de glutaminebehoeften toe in de periode na de training, waar de lokale immuunrespons kan concurreren om de beschikbaarheid van glutamine om spiercellen te stimuleren voor de opname van aminozuren en eiwitsynthese.
Voorladen cellen met glutamine kunnen mogelijk ook de "vertragingstijd" verminderen die wordt geassocieerd met leucine-activering van de eiwitsynthese. Als je dit nog niet doet, neem dan direct na de training 10-15 g glutamine of glutaminepeptiden. Omdat BCAA's een ander favoriet substraat zijn voor de synthese van spierglutamine en er ook is aangetoond dat ze de productie van spierglutamine verhogen (19-21), zijn BCAA's en leucine ook nuttig tijdens de pre-workoutperiode om de endogene glutamineproductie te helpen maximaliseren.
Noot van de redactie: het volledige Plamza / MAG-10-protocol, met zijn pre-training, training en post-training voeding, voldoet aan de aanbevelingen van de auteur voor het belasten van de spieren met BCAA's en leucine.
Onlangs werd ontdekt dat de consumptie van EAA's de expressie van zowel System A als System L aminozuurtransporters verhoogt.(9) Belangrijk is dat dit gebeurt op het 'posttranscriptieniveau', dat wil zeggen op het niveau van eiwitsynthese, waar bestaande mRNA's worden vertaald in eiwitten.
Vergelijk dit met "denovo"Eiwitexpressie - waarbij het 16 of meer uur kan duren om nieuwe mRNA's te synthetiseren, verwerken en transporteren - posttranscriptionele activering van eiwitsynthese kan in minuten tot uren plaatsvinden, waardoor cellen snel het niveau van bepaalde eiwitten kunnen verhogen wanneer dat nodig is.
Plots hebben we meer stimulans om een solide voedingsplan voor de peri-workout te hebben - EAA-inname tijdens de pre- en peri-workoutperioden loont grote post-workout door de expressie van aminozuurtransporters te verhogen, waardoor de cel wordt voorbereid op maximale opname van aminozuren en activering van eiwitsynthese.
Snel opneembare eiwitisolaten of hydrolysaten zoals Mag-10® Protein Pulsing Protocol ™ of Plazma ™ tijdens de pre- en peri-workout periodes zijn hier ideaal.
Insuline is het meest anabole hormoon in het lichaam. Naast het direct activeren van de eiwitsynthese, verhoogt insuline ook de translocatie van de aminozuurtransporteurs van Systeem A naar het celmembraan.(22)
Dit betekent dat insuline ervoor zorgt dat er meer System A-transporters op het celmembraan worden weergegeven, klaar om meer glutamine de cel in te drijven. Meer glutamine leidt tot meer celvolume, wat meer leucine de cel in drijft, wat uiteindelijk leidt tot meer eiwitsynthese.
Terwijl EAA's de uitdrukking van AA-transporters, het is het insulinesignaal waarmee ze op het celoppervlak kunnen worden weergegeven, klaar om nieuwe aminozuren de cel in te transporteren.
Dit is nog een andere reden waarom koolhydraten vóór en per training een goed idee zijn, tenzij u zich in de modus voor extreem vetverlies bevindt: insuline verhoogt de capaciteit voor cellulair aminozuurtransport.
Koolhydraten verhogen het insulinegehalte, maar bepaalde aminozuren kunnen ook worden gebruikt om de insulineafgifte te versterken. Glutamine is een krachtige activator van 'incretine'-hormonen, die insulineproducerende cellen in de alvleesklier gevoeliger maken voor glucose.(23) Glycine versterkt ook de afgifte van insuline via een ander mechanisme.
Hoewel alleen koolhydraten na de training het insulinegehalte verhogen, zal het combineren van deze insulinepotentiërende aminozuren met koolhydraten uw alvleesklier stimuleren voor nog meer insulineafgifte. Hoewel het goed is om het insulinegehalte meestal aan de lage kant te houden, maximaliseert een verhoogd insulinegehalte tijdens de peri-workoutperiode het aminozuurtransport, het celvolume en de eiwitsynthese, terwijl ook de afbraak van eiwitten wordt onderdrukt.
Het soort intense anaërobe training dat nodig is om veel kwaliteitsspieren op te bouwen, leidt tot een aanzienlijke lactaatproductie, waardoor de pH van de spieren daalt. Dit leidt tot vroege spiervermoeidheid en spierzwakte, maar ook bepaalde aminozuurtransporteurs, waaronder systeem A, worden geremd door een lage pH.(13)
Wanneer de spier-pH laag is, wordt de opname van aminozuren verminderd, wat de activering van mTOR van de eiwitsynthese onderdrukt.(24) Er is ook aangetoond dat remming van de lage pH van de aminozuurtransporteurs van systeem A de eiwitafbraak verhoogt.(25)
Dit is waar bèta-alanine binnenkomt. Verhoogde spiercarnosineniveaus werken als een natuurlijke zuurbuffer, waardoor de anaërobe drempel wordt verlengd door de afname van de spier-pH door training te beperken.
Beta-alanine heeft nog een andere belangrijke functie: om de eiwitsynthese te helpen behouden en deze sneller online te krijgen na intensieve training door de verzwakking van het aminozuurtransport te voorkomen.
Om het carnosinegehalte in de spieren te verhogen, dient u driemaal daags twee tabletten Beta-7 ™ in te nemen.
Tijdens intensieve trainingssessies wordt de eiwitsynthese verminderd en wordt de eiwitafbraak geactiveerd. Dit is onvermijdelijk voor elke harde trainingslift. De mate waarin we de katabole effecten van training kunnen minimaliseren en hoe sneller we kunnen terugkeren naar de 'anabole modus' tijdens de periode na de training, bepaalt uiteindelijk hoe efficiënt we zullen herstellen - en groeien.
De timing van macronutriënten is belangrijk, maar het is een middel om een doel te bereiken. Het celvolume is de belangrijkste motor van aminozuurtransport en eiwitsynthese. Door te begrijpen hoe aminozuurtransport plaatsvindt en hoe het wordt gereguleerd door celvolume, kunnen we meer leucine sneller in de verwoeste spiercellen krijgen, waardoor het anabole vuur wordt aangewakkerd en uiteindelijk leidt tot betere winsten.
De bovenstaande strategieën zijn effectief, praktisch en gebaseerd op het laatste wetenschappelijke onderzoek. Gebruik ze als sjabloon om uw trainingsvoortgang naar een hoger niveau te tillen.
Ik kijk uit naar uw vragen over de LiveSpill!



Niemand heeft nog op dit artikel gereageerd.